Global Patterns in Human Mitochodrial DNA and Y-Chromosome Variation Caused by Spatial Instabilityof the Local Cultural Processes
Vikrant Kumar, Banrida T. Langstieh, Komal V. Madhavi, Vegi M. Naidu, Hardeep Pal Singh, Silpak Biswas, Kumarasamy Thangaraj, Lalji Singh, B. Mohan Reddy
PLoS Genetics v. 2 April 2006
Synopsis: In most human societies, women traditionally move to their husband's home after marriage, and these societies are thus “patrilocal,” but in a few “matrilocal” societies, men move to their wife's home. These social customs are expected to influence the patterns of genetic variation. They should lead to a localization of male-specific Y-chromosomal variants and wide dispersal of female-specific mitochondrial DNA variants in patrilocal societies and vice versa in matrilocal societies. These predicted patterns have indeed been observed in previous studies of populations from Thailand. Indian societies, however, are endogamous, so marriage should always take place within a population, and these different patterns of genetic variation should not build up. The authors have now analyzed ten patrilocal and five matrilocal Indian populations, and find that there is indeed little difference between the patrilocal and matrilocal societies. The authors therefore conclude that patterns of genetic variation in humans are not universal, but depend on local cultural practices.
The authors assume that most human societies are patrilocal, but it seems that the jury is still out on this topic. See papers by Mark Shriver, Frank Marlowe, Wilder (cited in this paper).
Endogamy is quite strict in the Indian groups examined in this study. An equally important issue arises, namely, what determines variation in endogamy?
Friday, April 28, 2006
Wednesday, April 26, 2006
Linkage of trait and trait preference in butterfly
Linkage of butterfly mate preference and wing color preference cue at the genomic location of wingless
Marcus R. Kronforst, Laura G. Young, Durrell D. Kapan, Camille McNeely, Rachel J. O'Neill, and Lawrence E. Gilbert
PNAS April 25, 2006; 103:6575-6580
Abstract: Sexual isolation is a critical form of reproductive isolation in the early stages of animal speciation, yet little is known about the genetic basis of divergent mate preferences and preference cues in young species. Heliconius butterflies, well known for their diversity of wing color patterns, mate assortatively as a result of divergence in male preference for wing patterns. Here we show that the specific cue used by Heliconius cydno and Heliconius pachinus males to recognize conspecific females is the color of patches on the wings. In addition, male mate preference segregates with forewing color in hybrids, indicating a genetic association between the loci responsible for preference and preference cue. Quantitative trait locus mapping places a preference locus coincident with the locus that determines forewing color, which itself is perfectly linked to the wing patterning candidate gene, wingless. Furthermore, yellow-colored males of the polymorphic race H. cydno alithea prefer to court yellow females, indicating that wing color and color preference are controlled by loci that are located in an inversion or are pleiotropic effects of a single locus. Tight genetic associations between preference and preference cue, although rare, make divergence and speciation particularly likely because the effects of natural and sexual selection on one trait are transferred to the other, leading to the coordinated evolution of mate recognition. This effect of linkage on divergence is especially important in Heliconius because differentiation of wing color patterns in the genus has been driven and maintained by natural selection for Müllerian mimicry.
The last paragraph gives insight into the overall mechanism:
"Regardless of the specific nature of the association, the fact that wing color and color preference are very tightly linked genetically has important evolutionary consequences. Wing pattern diversification in the genus Heliconius has been driven by natural selection for Müllerian mimicry (33, 46, 47). Because wing color serves as a critical cue in mate recognition, and male preference for wing color is linked to color, natural selection on wing patterns that is imposed by predators will simultaneously drive the divergence of both mate recognition signal and signal preference. This scenario, which provides a direct link between disruptive natural selection and divergence at loci mediating conspecific recognition and mate choice, facilitates the rapid evolution of sexual isolation. Furthermore, the association between color and preference limits recombination between the two components of mate recognition when species and color pattern races hybridize, a common phenomenon in the genus Heliconius, thus allowing phenotypic differentiation to persist in the face of hybridization. Given the effects of this association on the origin and maintenance of diversity, it has undoubtedly played a significant role in facilitating the explosive adaptive radiation of Heliconius butterflies."
Marcus R. Kronforst, Laura G. Young, Durrell D. Kapan, Camille McNeely, Rachel J. O'Neill, and Lawrence E. Gilbert
PNAS April 25, 2006; 103:6575-6580
Abstract: Sexual isolation is a critical form of reproductive isolation in the early stages of animal speciation, yet little is known about the genetic basis of divergent mate preferences and preference cues in young species. Heliconius butterflies, well known for their diversity of wing color patterns, mate assortatively as a result of divergence in male preference for wing patterns. Here we show that the specific cue used by Heliconius cydno and Heliconius pachinus males to recognize conspecific females is the color of patches on the wings. In addition, male mate preference segregates with forewing color in hybrids, indicating a genetic association between the loci responsible for preference and preference cue. Quantitative trait locus mapping places a preference locus coincident with the locus that determines forewing color, which itself is perfectly linked to the wing patterning candidate gene, wingless. Furthermore, yellow-colored males of the polymorphic race H. cydno alithea prefer to court yellow females, indicating that wing color and color preference are controlled by loci that are located in an inversion or are pleiotropic effects of a single locus. Tight genetic associations between preference and preference cue, although rare, make divergence and speciation particularly likely because the effects of natural and sexual selection on one trait are transferred to the other, leading to the coordinated evolution of mate recognition. This effect of linkage on divergence is especially important in Heliconius because differentiation of wing color patterns in the genus has been driven and maintained by natural selection for Müllerian mimicry.
The last paragraph gives insight into the overall mechanism:
"Regardless of the specific nature of the association, the fact that wing color and color preference are very tightly linked genetically has important evolutionary consequences. Wing pattern diversification in the genus Heliconius has been driven by natural selection for Müllerian mimicry (33, 46, 47). Because wing color serves as a critical cue in mate recognition, and male preference for wing color is linked to color, natural selection on wing patterns that is imposed by predators will simultaneously drive the divergence of both mate recognition signal and signal preference. This scenario, which provides a direct link between disruptive natural selection and divergence at loci mediating conspecific recognition and mate choice, facilitates the rapid evolution of sexual isolation. Furthermore, the association between color and preference limits recombination between the two components of mate recognition when species and color pattern races hybridize, a common phenomenon in the genus Heliconius, thus allowing phenotypic differentiation to persist in the face of hybridization. Given the effects of this association on the origin and maintenance of diversity, it has undoubtedly played a significant role in facilitating the explosive adaptive radiation of Heliconius butterflies."
Tuesday, April 25, 2006
Mice that can run and run and run
I thought this paper in PLoS Biology from a couple years back was pretty interesting/funny given that I have some interest in the evolutionary ecology and genetics of muscle physiology and sports.
Here's a video of the transgenic mouse that keeps on running, while the wild-type is getting exhausted. By the way, exhaustion is defined as being "unable to avoid repetitive electrical shocks."... poor mice!
Regulation of Muscle Fiber Type and Running Endurance by PPARδ
Yong-Xu Wang, Chun-Li Zhang, Ruth T. Yu, Helen K. Cho, Michael C. Nelson, Corinne R. Bayuga-Ocampo, Jungyeob Ham, Heonjoong Kang, Ronald M. Evans*
Abstract: Endurance exercise training can promote an adaptive muscle fiber transformation and an increase of mitochondrial biogenesis by triggering scripted changes in gene expression. However, no transcription factor has yet been identified that can direct this process. We describe the engineering of a mouse capable of continuous running of up to twice the distance of a wild-type littermate. This was achieved by targeted expression of an activated form of peroxisome proliferator-activated receptor δ (PPARδ) in skeletal muscle, which induces a switch to form increased numbers of type I muscle fibers. Treatment of wild-type mice with PPARδ agonist elicits a similar type I fiber gene expression profile in muscle. Moreover, these genetically generated fibers confer resistance to obesity with improved metabolic profiles, even in the absence of exercise. These results demonstrate that complex physiologic properties such as fatigue, endurance, and running capacity can be molecularly analyzed and manipulated.
A few genes implicated in type I and type II muscle fibers, or associated with endurance vs. sprint performance have been found in humans but those are not mentioned in this paper. I wonder if anyone has looked at PPAR in humans.
Here's a video of the transgenic mouse that keeps on running, while the wild-type is getting exhausted. By the way, exhaustion is defined as being "unable to avoid repetitive electrical shocks."... poor mice!
Regulation of Muscle Fiber Type and Running Endurance by PPARδ
Yong-Xu Wang, Chun-Li Zhang, Ruth T. Yu, Helen K. Cho, Michael C. Nelson, Corinne R. Bayuga-Ocampo, Jungyeob Ham, Heonjoong Kang, Ronald M. Evans*
Abstract: Endurance exercise training can promote an adaptive muscle fiber transformation and an increase of mitochondrial biogenesis by triggering scripted changes in gene expression. However, no transcription factor has yet been identified that can direct this process. We describe the engineering of a mouse capable of continuous running of up to twice the distance of a wild-type littermate. This was achieved by targeted expression of an activated form of peroxisome proliferator-activated receptor δ (PPARδ) in skeletal muscle, which induces a switch to form increased numbers of type I muscle fibers. Treatment of wild-type mice with PPARδ agonist elicits a similar type I fiber gene expression profile in muscle. Moreover, these genetically generated fibers confer resistance to obesity with improved metabolic profiles, even in the absence of exercise. These results demonstrate that complex physiologic properties such as fatigue, endurance, and running capacity can be molecularly analyzed and manipulated.
A few genes implicated in type I and type II muscle fibers, or associated with endurance vs. sprint performance have been found in humans but those are not mentioned in this paper. I wonder if anyone has looked at PPAR in humans.
Monday, April 24, 2006
Ten Simple Rules for Getting Published
by Philip E. Bourne
PLOS Computational Biology v. Issue 5, October 2005
"The student council (http://www.iscbsc.org/) of the International Society for Computational Biology asked me to present my thoughts on getting published in the field of computational biology at the Intelligent Systems in Molecular Biology conference held in Detroit in late June of 2005. Close to 200 bright young souls (and a few not so young) crammed into a small room for what proved to be a wonderful interchange among a group of whom approximately one-half had yet to publish their first paper. The advice I gave that day I have modified and present as ten rules for getting published.
Rule 1: Read many papers, and learn from both the good and the bad work of others.
It is never too early to become a critic. Journal clubs, where you critique a paper as a group, are excellent for having this kind of dialogue. Reading at least two papers a day in detail (not just in your area of research) and thinking about their quality will also help. Being well read has another potential major benefit—it facilitates a more objective view of one's own work. It is too easy after many late nights spent in front of a computer screen and/or laboratory bench to convince yourself that your work is the best invention since sliced bread. More than likely it is not, and your mentor is prone to falling into the same trap, hence rule 2.
Rule 2: The more objective you can be about your work, the better that work will ultimately become.
Alas, some scientists will never be objective about their own work, and will never make the best scientists—learn objectivity early, the editors and reviewers have.
Rule 3: Good editors and reviewers will be objective about your work.
The quality of the editorial board is an early indicator of the review process. Look at the masthead of the journal in which you plan to publish. Outstanding editors demand and get outstanding reviews. Put your energy into improving the quality of the manuscript before submission. Ideally, the reviews will improve your paper. But they will not get to imparting that advice if there are fundamental flaws.
Rule 4: If you do not write well in the English language, take lessons early; it will be invaluable later.
This is not just about grammar, but more importantly comprehension. The best papers are those in which complex ideas are expressed in a way that those who are less than immersed in the field can understand. Have you noticed that the most renowned scientists often give the most logical and simply stated yet stimulating lectures? This extends to their written work as well. Note that writing clearly is valuable, even if your ultimate career does not hinge on producing good scientific papers in English language journals. Submitted papers that are not clearly written in good English, unless the science is truly outstanding, are often rejected or at best slow to publish since they require extensive copyediting.
Rule 5: Learn to live with rejection.
A failure to be objective can make rejection harder to take, and you will be rejected. Scientific careers are full of rejection, even for the best scientists. The correct response to a paper being rejected or requiring major revision is to listen to the reviewers and respond in an objective, not subjective, manner. Reviews reflect how your paper is being judged—learn to live with it. If reviewers are unanimous about the poor quality of the paper, move on—in virtually all cases, they are right. If they request a major revision, do it and address every point they raise both in your cover letter and through obvious revisions to the text. Multiple rounds of revision are painful for all those concerned and slow the publishing process.
Rule 6: The ingredients of good science are obvious—novelty of research topic, comprehensive coverage of the relevant literature, good data, good analysis including strong statistical support, and a thought-provoking discussion. The ingredients of good science reporting are obvious—good organization, the appropriate use of tables and figures, the right length, writing to the intended audience—do not ignore the obvious.
Be objective about these ingredients when you review the first draft, and do not rely on your mentor. Get a candid opinion by having the paper read by colleagues without a vested interest in the work, including those not directly involved in the topic area.
Rule 7: Start writing the paper the day you have the idea of what questions to pursue.
Some would argue that this places too much emphasis on publishing, but it could also be argued that it helps define scope and facilitates hypothesis-driven science. The temptation of novice authors is to try to include everything they know in a paper. Your thesis is/was your kitchen sink. Your papers should be concise, and impart as much information as possible in the least number of words. Be familiar with the guide to authors and follow it, the editors and reviewers do. Maintain a good bibliographic database as you go, and read the papers in it.
Rule 8: Become a reviewer early in your career.
Reviewing other papers will help you write better papers. To start, work with your mentors; have them give you papers they are reviewing and do the first cut at the review (most mentors will be happy to do this). Then, go through the final review that gets sent in by your mentor, and where allowed, as is true of this journal, look at the reviews others have written. This will provide an important perspective on the quality of your reviews and, hopefully, allow you to see your own work in a more objective way. You will also come to understand the review process and the quality of reviews, which is an important ingredient in deciding where to send your paper.
Rule 9: Decide early on where to try to publish your paper.
This will define the form and level of detail and assumed novelty of the work you are doing. Many journals have a presubmission enquiry system available—use it. Even before the paper is written, get a sense of the novelty of the work, and whether a specific journal will be interested.
Rule 10: Quality is everything.
It is better to publish one paper in a quality journal than multiple papers in lesser journals. Increasingly, it is harder to hide the impact of your papers; tools like Google Scholar and the ISI Web of Science are being used by tenure committees and employers to define metrics for the quality of your work. It used to be that just the journal name was used as a metric. In the digital world, everyone knows if a paper has little impact. Try to publish in journals that have high impact factors; chances are your paper will have high impact, too, if accepted.
When you are long gone, your scientific legacy is, in large part, the literature you left behind and the impact it represents. I hope these ten simple rules can help you leave behind something future generations of scientists will admire. "
PLOS Computational Biology v. Issue 5, October 2005
"The student council (http://www.iscbsc.org/) of the International Society for Computational Biology asked me to present my thoughts on getting published in the field of computational biology at the Intelligent Systems in Molecular Biology conference held in Detroit in late June of 2005. Close to 200 bright young souls (and a few not so young) crammed into a small room for what proved to be a wonderful interchange among a group of whom approximately one-half had yet to publish their first paper. The advice I gave that day I have modified and present as ten rules for getting published.
Rule 1: Read many papers, and learn from both the good and the bad work of others.
It is never too early to become a critic. Journal clubs, where you critique a paper as a group, are excellent for having this kind of dialogue. Reading at least two papers a day in detail (not just in your area of research) and thinking about their quality will also help. Being well read has another potential major benefit—it facilitates a more objective view of one's own work. It is too easy after many late nights spent in front of a computer screen and/or laboratory bench to convince yourself that your work is the best invention since sliced bread. More than likely it is not, and your mentor is prone to falling into the same trap, hence rule 2.
Rule 2: The more objective you can be about your work, the better that work will ultimately become.
Alas, some scientists will never be objective about their own work, and will never make the best scientists—learn objectivity early, the editors and reviewers have.
Rule 3: Good editors and reviewers will be objective about your work.
The quality of the editorial board is an early indicator of the review process. Look at the masthead of the journal in which you plan to publish. Outstanding editors demand and get outstanding reviews. Put your energy into improving the quality of the manuscript before submission. Ideally, the reviews will improve your paper. But they will not get to imparting that advice if there are fundamental flaws.
Rule 4: If you do not write well in the English language, take lessons early; it will be invaluable later.
This is not just about grammar, but more importantly comprehension. The best papers are those in which complex ideas are expressed in a way that those who are less than immersed in the field can understand. Have you noticed that the most renowned scientists often give the most logical and simply stated yet stimulating lectures? This extends to their written work as well. Note that writing clearly is valuable, even if your ultimate career does not hinge on producing good scientific papers in English language journals. Submitted papers that are not clearly written in good English, unless the science is truly outstanding, are often rejected or at best slow to publish since they require extensive copyediting.
Rule 5: Learn to live with rejection.
A failure to be objective can make rejection harder to take, and you will be rejected. Scientific careers are full of rejection, even for the best scientists. The correct response to a paper being rejected or requiring major revision is to listen to the reviewers and respond in an objective, not subjective, manner. Reviews reflect how your paper is being judged—learn to live with it. If reviewers are unanimous about the poor quality of the paper, move on—in virtually all cases, they are right. If they request a major revision, do it and address every point they raise both in your cover letter and through obvious revisions to the text. Multiple rounds of revision are painful for all those concerned and slow the publishing process.
Rule 6: The ingredients of good science are obvious—novelty of research topic, comprehensive coverage of the relevant literature, good data, good analysis including strong statistical support, and a thought-provoking discussion. The ingredients of good science reporting are obvious—good organization, the appropriate use of tables and figures, the right length, writing to the intended audience—do not ignore the obvious.
Be objective about these ingredients when you review the first draft, and do not rely on your mentor. Get a candid opinion by having the paper read by colleagues without a vested interest in the work, including those not directly involved in the topic area.
Rule 7: Start writing the paper the day you have the idea of what questions to pursue.
Some would argue that this places too much emphasis on publishing, but it could also be argued that it helps define scope and facilitates hypothesis-driven science. The temptation of novice authors is to try to include everything they know in a paper. Your thesis is/was your kitchen sink. Your papers should be concise, and impart as much information as possible in the least number of words. Be familiar with the guide to authors and follow it, the editors and reviewers do. Maintain a good bibliographic database as you go, and read the papers in it.
Rule 8: Become a reviewer early in your career.
Reviewing other papers will help you write better papers. To start, work with your mentors; have them give you papers they are reviewing and do the first cut at the review (most mentors will be happy to do this). Then, go through the final review that gets sent in by your mentor, and where allowed, as is true of this journal, look at the reviews others have written. This will provide an important perspective on the quality of your reviews and, hopefully, allow you to see your own work in a more objective way. You will also come to understand the review process and the quality of reviews, which is an important ingredient in deciding where to send your paper.
Rule 9: Decide early on where to try to publish your paper.
This will define the form and level of detail and assumed novelty of the work you are doing. Many journals have a presubmission enquiry system available—use it. Even before the paper is written, get a sense of the novelty of the work, and whether a specific journal will be interested.
Rule 10: Quality is everything.
It is better to publish one paper in a quality journal than multiple papers in lesser journals. Increasingly, it is harder to hide the impact of your papers; tools like Google Scholar and the ISI Web of Science are being used by tenure committees and employers to define metrics for the quality of your work. It used to be that just the journal name was used as a metric. In the digital world, everyone knows if a paper has little impact. Try to publish in journals that have high impact factors; chances are your paper will have high impact, too, if accepted.
When you are long gone, your scientific legacy is, in large part, the literature you left behind and the impact it represents. I hope these ten simple rules can help you leave behind something future generations of scientists will admire. "
Thursday, April 20, 2006
Estimates of mutation rates in humans
In this paper by J. Shea in the current issue of Evol. Anthr. mentions:
"The chronological basis for (pre)historical interpretations of living human genetic variation (thatis, mutation rates) are apparently rooted in archaeological estimates for biogeographic separation of human populations. These estimates are controversial, to say the least"
The paper is about a conference called "Rethinking the Human Revolution", and as usual talks about the need for more inter-sub (and subsub)disciplinary exchange and understanding...like between archaeologists and geneticits, as evidenced by the uncertainty mentioned in the above quote, and "between European and Near Eastern archeologists working in a culture-historical research tradition and, again largely, Anglo-American archaeologists working in an evolutionary ecological one."
"The chronological basis for (pre)historical interpretations of living human genetic variation (thatis, mutation rates) are apparently rooted in archaeological estimates for biogeographic separation of human populations. These estimates are controversial, to say the least"
The paper is about a conference called "Rethinking the Human Revolution", and as usual talks about the need for more inter-sub (and subsub)disciplinary exchange and understanding...like between archaeologists and geneticits, as evidenced by the uncertainty mentioned in the above quote, and "between European and Near Eastern archeologists working in a culture-historical research tradition and, again largely, Anglo-American archaeologists working in an evolutionary ecological one."
Wednesday, April 19, 2006
The Thrify Genotype hypothesis, food shortages, and the etiology of diabetes

Evolutionary perspectives on diabetes and other chronic diseases have primarily focused on the thrifty gene hypothesis. This perspective rests on the assumption that forager populations experienced more food shortages than agricultural populations. This paper, as well as others (cited in this one) present cross-cultural evidence that there is no difference between foragers and agricultural groups in the prevalence and severity of food shortages. (more below the abstract)
Exploring the thrifty genotype's food-shortage assumptions: A cross-cultural comparison of ethnographic accounts of food security among foraging and agricultural societies
Daniel C. Benyshek, James T. Watson: American Journal of Physical Anthropology, Early view: Feb 2006
Abstract:The "thrifty genotype hypothesis" has become firmly entrenched as one of the orienting concepts in biomedical anthropology, since first being proposed by Neel ([1962] Am. J. Hum. Genet. 14:353-362) over 40 years ago. Its influence on inquiries into the evolutionary origins of diabetes, lactose tolerance, and other metabolic disorders can hardly be underestimated, as evidenced by its continued citation in many top scientific and medical journals. However, its fundamental assumption, that foragers are more likely to experience regular and severe food shortages than sedentary agriculturalists, remains largely untested. The present report tests this assumption by making a cross-cultural statistical comparison of the quantity of available food and the frequency and extent of food shortages among 94 foraging and agricultural societies as reported in the ethnographic record. Our results indicate that there is no statistical difference in the quantity of available food, or the frequency or extent of food shortages in these reports between preindustrial foragers, recent foragers, and agriculturalists. The findings presented here add to a growing literature that calls into question assumptions about forager food insecurity and nutritional status in general, and ultimately, the very foundation of the thrifty genotype hypothesis: the presumed food shortages that selected for a "thrifty" metabolism in past foraging populations.
This paper does not consider food quality (fat vs. protein vs. carbohydrates etc...). Why is so important for women to have so many fat stores to begin pregnancy? Why not pay as you go, if food shortages (fat specifically) are not a problem?
Finally, the authors briefly discuss alternative hypotheses (fetal origin model) for the etiology of diabetes.
Tuesday, April 18, 2006
Gene expression levels of HoxA5, Gpc4 and Tbx15 correlated with BMI and/or WHR
This paper in PNAS reports findings that expression levels of several genes in human adipose tissue are predictive of BMI and/or WHR.
Evidence for a role of developmental genes in the origin of obesity and body fat distribution
Stephane Gesta, C. Ronald Kahn et al.
Published online before print April 14, 2006
PNAS
Abstract: Obesity, especially central obesity, is a hereditable trait associated with a high risk for development of diabetes and metabolic disorders. Combined gene expression analysis of adipocyte- and preadipocyte-containing fractions from intraabdominal and subcutaneous adipose tissue of mice revealed coordinated depot-specific differences in expression of multiple genes involved in embryonic development and pattern specification. These differences were intrinsic and persisted during in vitro culture and differentiation. Similar depot-specific differences in expression of developmental genes were observed in human subcutaneous versus visceral adipose tissue. Furthermore, in humans, several genes exhibited changes in expression that correlated closely with body mass index and/or waist/hip ratio. Together, these data suggest that genetically programmed developmental differences in adipocytes and their precursors in different regions of the body play an important role in obesity, body fat distribution, and potential functional differences between internal and subcutaneous adipose tissue.
Evidence for a role of developmental genes in the origin of obesity and body fat distribution
Stephane Gesta, C. Ronald Kahn et al.
Published online before print April 14, 2006
PNAS
Abstract: Obesity, especially central obesity, is a hereditable trait associated with a high risk for development of diabetes and metabolic disorders. Combined gene expression analysis of adipocyte- and preadipocyte-containing fractions from intraabdominal and subcutaneous adipose tissue of mice revealed coordinated depot-specific differences in expression of multiple genes involved in embryonic development and pattern specification. These differences were intrinsic and persisted during in vitro culture and differentiation. Similar depot-specific differences in expression of developmental genes were observed in human subcutaneous versus visceral adipose tissue. Furthermore, in humans, several genes exhibited changes in expression that correlated closely with body mass index and/or waist/hip ratio. Together, these data suggest that genetically programmed developmental differences in adipocytes and their precursors in different regions of the body play an important role in obesity, body fat distribution, and potential functional differences between internal and subcutaneous adipose tissue.
Thursday, April 13, 2006
More LD in population isolates
Published online: Nature Genetics, 2 April 2006;
Magnitude and distribution of linkage disequilibrium in population isolates and implications for genome-wide association studies
Susan Service et al
Abstract: The genome-wide distribution of linkage disequilibrium (LD) determines the strategy for selecting markers for association studies, but it varies between populations. We assayed LD in large samples (200 individuals) from each of 11 well-described population isolates and an outbred European-derived sample, using SNP markers spaced across chromosome 22. Most isolates show substantially higher levels of LD than the outbred sample and many fewer regions of very low LD (termed 'holes'). Young isolates known to have had relatively few founders show particularly extensive LD with very few holes; these populations offer substantial advantages for genome-wide association mapping.
Here is the supplementary table containing info. on the populations that were examined.
The Kuusamo population of Finland showed the highest amount of LD, with the fewest holes (regions of low LD). The authors attribut this to a recent founding by a few individuals, population bottleneck and rapid expansion.
Magnitude and distribution of linkage disequilibrium in population isolates and implications for genome-wide association studies
Susan Service et al
Abstract: The genome-wide distribution of linkage disequilibrium (LD) determines the strategy for selecting markers for association studies, but it varies between populations. We assayed LD in large samples (200 individuals) from each of 11 well-described population isolates and an outbred European-derived sample, using SNP markers spaced across chromosome 22. Most isolates show substantially higher levels of LD than the outbred sample and many fewer regions of very low LD (termed 'holes'). Young isolates known to have had relatively few founders show particularly extensive LD with very few holes; these populations offer substantial advantages for genome-wide association mapping.
Here is the supplementary table containing info. on the populations that were examined.
The Kuusamo population of Finland showed the highest amount of LD, with the fewest holes (regions of low LD). The authors attribut this to a recent founding by a few individuals, population bottleneck and rapid expansion.
Wednesday, April 12, 2006
Sex differences in senescence reconsidered
A reassessment of sexual dimorphism in human senescence: Theory, evidence, and causation
Brent M. Graves, Mac Strand, Alec R. Lindsay, AmJHumBio (2006) 18:161- 168
Abstract: Age-specific mortality rates of men are higher than those of women, and men have shorter average life spans than women. This has been interpreted as evidence of sexual dimorphism in rates of senescence. However, because mortality can be caused by numerous factors in addition to senescence, higher mortality rates do not necessarily indicate more rapid senescence. In this paper, we (1) emphasize the necessity of decoupling mortality and senescence when considering sexual dimorphism in senescence, (2) present a theoretical framework for the hypothesis that selection affects senescence in human males and females differently due to different life history characteristics, (3) consider phenotypic evidence from the literature that human males show a later onset of senescence than human females, despite exhibiting higher mortality rates, and (4) discuss the potential roles of mutation accumulation and antagonistic pleiotropy in the evolution of sexual dimorphism in senescence. Am. J. Hum. Biol. 18:161-168, 2006. © 2006 Wiley-Liss, Inc.
Who gets older and weaker faster? Men or women? Depending on what you're looking at, the evidence seems very mixed. This paper calls for the need to seperate mortality and senescence. The authors state that even though male life span is shorter, men actually senesce slower. They cite evidence from a variety of studies. This literature review is far from complete. I have reviewed this literature (sports physiology, physical activity levels, gerontology) in the last few months, and have had a hard time finding consistent sex differences in senescence.
The authors discuss the evolutionary mechanisms underlying sexual dimorphism in senescence and conclude that it must evolve via the antagonistic pleiotropy process, rather than the mutation accumulation process. They state that:
"This conclusion illustrates our perspective that the evolution of sexual dimorphism in senescence should be considered in different ways from the evolution of senescence itself."
Brent M. Graves, Mac Strand, Alec R. Lindsay, AmJHumBio (2006) 18:161- 168
Abstract: Age-specific mortality rates of men are higher than those of women, and men have shorter average life spans than women. This has been interpreted as evidence of sexual dimorphism in rates of senescence. However, because mortality can be caused by numerous factors in addition to senescence, higher mortality rates do not necessarily indicate more rapid senescence. In this paper, we (1) emphasize the necessity of decoupling mortality and senescence when considering sexual dimorphism in senescence, (2) present a theoretical framework for the hypothesis that selection affects senescence in human males and females differently due to different life history characteristics, (3) consider phenotypic evidence from the literature that human males show a later onset of senescence than human females, despite exhibiting higher mortality rates, and (4) discuss the potential roles of mutation accumulation and antagonistic pleiotropy in the evolution of sexual dimorphism in senescence. Am. J. Hum. Biol. 18:161-168, 2006. © 2006 Wiley-Liss, Inc.
Who gets older and weaker faster? Men or women? Depending on what you're looking at, the evidence seems very mixed. This paper calls for the need to seperate mortality and senescence. The authors state that even though male life span is shorter, men actually senesce slower. They cite evidence from a variety of studies. This literature review is far from complete. I have reviewed this literature (sports physiology, physical activity levels, gerontology) in the last few months, and have had a hard time finding consistent sex differences in senescence.
The authors discuss the evolutionary mechanisms underlying sexual dimorphism in senescence and conclude that it must evolve via the antagonistic pleiotropy process, rather than the mutation accumulation process. They state that:
"This conclusion illustrates our perspective that the evolution of sexual dimorphism in senescence should be considered in different ways from the evolution of senescence itself."
Friday, April 07, 2006
Punishment - an essential ingredient in successful groups
Science 7 April 2006 v312:108-111
The Competitive Advantage of Sanctioning Institutions
Özgür Gürerk, Bernd Irlenbusch, Bettina Rockenbach
Abstract: Understanding the fundamental patterns and determinants of human cooperation and the maintenance of social order in human societies is a challenge across disciplines. The existing empirical evidence for the higher levels of cooperation when altruistic punishment is present versus when it is absent systematically ignores the institutional competition inherent in human societies. Whether punishment would be deliberately adopted and would similarly enhance cooperation when directly competing with nonpunishment institutions is highly controversial in light of recent findings on the detrimental effects of punishment. We show experimentally that a sanctioning institution is the undisputed winner in a competition with a sanction-free institution. Despite initial aversion, the entire population migrates successively to the sanctioning institution and strongly cooperates, whereas the sanction-free society becomes fully depopulated. The findings demonstrate the competitive advantage of sanctioning institutions and exemplify the emergence and manifestation of social order driven by institutional selection.
The Competitive Advantage of Sanctioning Institutions
Özgür Gürerk, Bernd Irlenbusch, Bettina Rockenbach
Abstract: Understanding the fundamental patterns and determinants of human cooperation and the maintenance of social order in human societies is a challenge across disciplines. The existing empirical evidence for the higher levels of cooperation when altruistic punishment is present versus when it is absent systematically ignores the institutional competition inherent in human societies. Whether punishment would be deliberately adopted and would similarly enhance cooperation when directly competing with nonpunishment institutions is highly controversial in light of recent findings on the detrimental effects of punishment. We show experimentally that a sanctioning institution is the undisputed winner in a competition with a sanction-free institution. Despite initial aversion, the entire population migrates successively to the sanctioning institution and strongly cooperates, whereas the sanction-free society becomes fully depopulated. The findings demonstrate the competitive advantage of sanctioning institutions and exemplify the emergence and manifestation of social order driven by institutional selection.
From water to land 375 million years ago
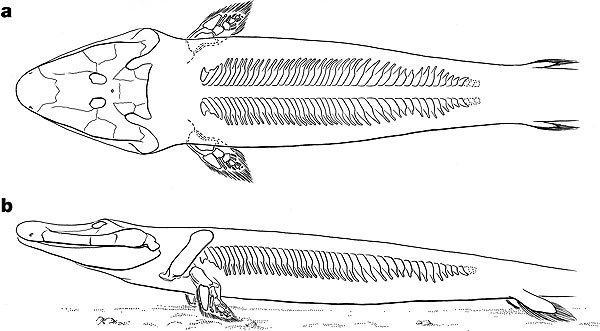
Nature has a paper describing fossil evidence for an ancient transitional form between fish and tetrapods, found in northern Canada.
From the abstract: "The morphological features and geological setting of this new animal are suggestive of life in shallow-water, marginal and subaerial habitats."
Edward B. Daeschler, Neil H. Shubin and Farish A
Nature 440: 757-763 (April 6, 2006)
Thursday, April 06, 2006
Cockroach Grouping
An interesting post at John Hawks' weblog on cockroach group behavior, giving us interesting/funny insight into how social behavior "works".
link to the PNAS abstract
link to the PNAS abstract
"Green Beard" reconsidered (once again)
A new paper in Nature (440:663-666), by Vincent Jansen and Minus van Baalen re-examines the plausibility of a green beard type of mechanism for altruism between individuals who have similar phenotypic "tags". The authors use simulations of Prisoner's Dilemma games in a spatial setting.
Abstract: The evolution of altruism, a behaviour that benefits others at one's own fitness expense, poses a darwinian paradox. The paradox is resolved if many interactions are with related individuals so that the benefits of altruism are reaped by copies of the altruistic gene in other individuals1, a mechanism called kin selection2. However, recognition of altruists could provide an alternative route towards the evolution of altruism1, 3, 4, 5. Arguably the simplest recognition system is a conspicuous, heritable tag, such as a green beard1, 3. Despite the fact that such genes have been reported6, 7, 8, the 'green beard effect'3 has often been dismissed because it is unlikely that a single gene can code for altruism and a recognizable tag1, 3, 9. Here we model the green beard effect and find that if recognition and altruism are always inherited together, the dynamics are highly unstable, leading to the loss of altruism. In contrast, if the effect is caused by loosely coupled separate genes, altruism is facilitated through beard chromodynamics in which many beard colours co-occur. This allows altruism to persist even in weakly structured populations and implies that the green beard effect, in the form of a fluid association of altruistic traits with a recognition tag, can be much more prevalent than hitherto assumed.
The last few sentences are the most interesting:
"Our analysis leads to the testable hypothesis that diversity in recognition tags inversely correlates with average relatedness. This suggests that relatively easily observed tag diversity can serve as an indicator for the nature of the underlying social interactions. Third, to detect the green beard effect one should look for cases where tag and trait can dynamically associate8, 16. A tag that functions as a green beard in one instance need not be associated with altruism in another population or at another instance in time, which obviously has consequences for our capacity to detect green beards."
One can imagine that this would have important implications in terms of the evolution of cooperation, phenotype matching, phenotypic diversity, and reproductive bahavior.
Abstract: The evolution of altruism, a behaviour that benefits others at one's own fitness expense, poses a darwinian paradox. The paradox is resolved if many interactions are with related individuals so that the benefits of altruism are reaped by copies of the altruistic gene in other individuals1, a mechanism called kin selection2. However, recognition of altruists could provide an alternative route towards the evolution of altruism1, 3, 4, 5. Arguably the simplest recognition system is a conspicuous, heritable tag, such as a green beard1, 3. Despite the fact that such genes have been reported6, 7, 8, the 'green beard effect'3 has often been dismissed because it is unlikely that a single gene can code for altruism and a recognizable tag1, 3, 9. Here we model the green beard effect and find that if recognition and altruism are always inherited together, the dynamics are highly unstable, leading to the loss of altruism. In contrast, if the effect is caused by loosely coupled separate genes, altruism is facilitated through beard chromodynamics in which many beard colours co-occur. This allows altruism to persist even in weakly structured populations and implies that the green beard effect, in the form of a fluid association of altruistic traits with a recognition tag, can be much more prevalent than hitherto assumed.
The last few sentences are the most interesting:
"Our analysis leads to the testable hypothesis that diversity in recognition tags inversely correlates with average relatedness. This suggests that relatively easily observed tag diversity can serve as an indicator for the nature of the underlying social interactions. Third, to detect the green beard effect one should look for cases where tag and trait can dynamically associate8, 16. A tag that functions as a green beard in one instance need not be associated with altruism in another population or at another instance in time, which obviously has consequences for our capacity to detect green beards."
One can imagine that this would have important implications in terms of the evolution of cooperation, phenotype matching, phenotypic diversity, and reproductive bahavior.
Subscribe to:
Posts (Atom)

